
周琦实验室在Nat. Commun. & Mol. Biol. Evol.同时发表两篇文章系统重构动物手指/脚趾的演化发育脉络
人灵巧的手指、鼹鼠挖洞的爪子、马奔跑用的蹄、鲸游泳用鳍、蝙蝠和鸟飞翔的翅膀等等源自同一套原始的祖先发育蓝图。四肢在数亿年演化中被空气,海洋,土地等截然不同的自然环境选择反复雕琢,承担不可或缺的功能,也形成了我们最熟悉的肢体的多样性。达尔文将这种同源性视作演化最有力的证据之一。然而,围绕这个经典问题,一场跨越近两百年的争论至今未歇:鸟类翅膀只有三根甚至一根手指,到底对应着祖先的哪几根手指?这个看似钻脑洞的问题,却是研究细胞器官水平同源性的重要范式。尽管现代基因组学的发展使得我们可以非常清晰的确定基因在物种间的同源性,但是一旦上升到细胞和器官水平,却远非那么简单。因为在细胞和器官层面,我们在比较的是几百个基因所形成的决定细胞或器官身份的网络。那套决定拇指是拇指、小指是小指的基因调控网络,在脊椎动物从鱼鳍演化出四肢的漫长历程中,究竟多少作为核心工具基因(toolbox gene)被恒久远地保留?又有哪些经历了我们所谓空气,海洋,土地等的重塑发生了物种特异的变化?
近日,我院周琦教授课题组Nature Communications和Molecular Biology and Evolution期刊同时发表两篇研究论文,通过比较转录组学的全新视角,从宏观演化框架到具体物种案例,系统阐释了四足动物指/趾的身份决定、退化与适应性演化的分子机制。前者全景勾勒了自有颌类脊椎动物祖先以来指/趾身份基因网络的演化图景,后者聚焦平胸鸟类(鸸鹋与鸵鸟)的具体案例,拆解指/趾退化和适应的不同分子路径。


鸟类翼指的分子特征是1-3-4
大多数现生鸟类有三根翼指和两到四根脚趾。化石证据表明,从恐龙到鸟类的过渡中,前肢指数量由五根减至三根。一种基于化石证据的推测是:鸟类丢失了无名指和小指,保留了大拇指、食指和中指(即1、2、3指)。因此鸟类手指与恐龙的手指如此连续的化石线索一直以来被认为是“鸟类就是孑遗恐龙”的重要证据。然而,鸡胚的发育生物学和转录组数据却支持翅膀三根手指分别为拇指、中指和无名指(1、3、4指)——这便是框架位移(frameshift)假说:手指的位置身份和发育身份发生了错位,原本应长在第二位的指头,却呈现第一指的分子特征。
为了彻底解决这一争议,研究团队收集了鸡、鸵鸟、鸸鹋、暹罗鳄、中华鳖和墨西哥钝口螈共六个物种的肢芽和每一根发育过程中指/趾的转录组数据,横跨三个发育阶段,并整合了已发表的绿安乐蜥和小鼠数据——这是迄今为止最大规模的四足动物手指/趾比较转录组数据集。
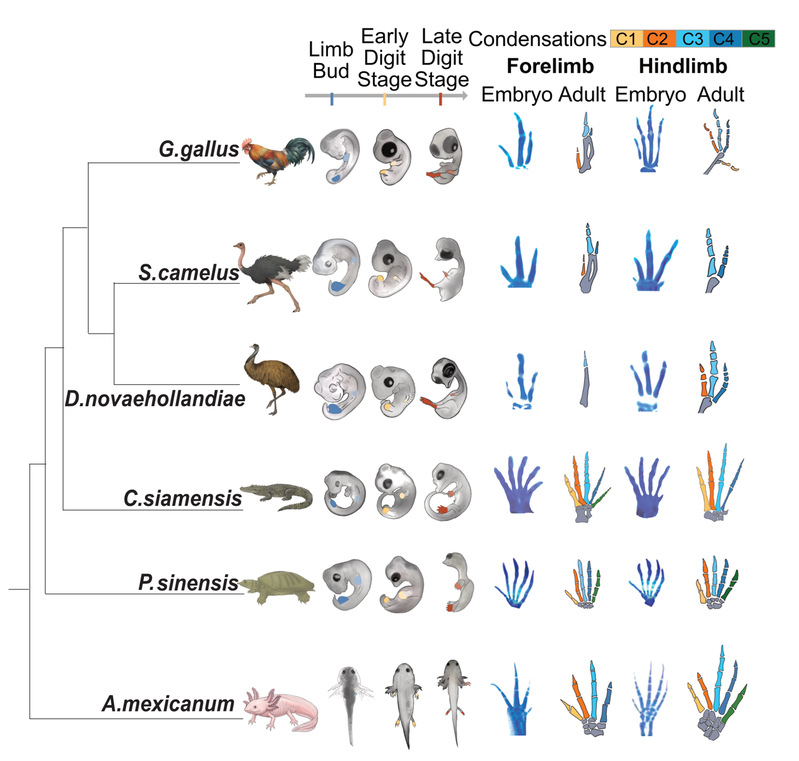
▲ 图一、研究涵盖的六个四足动物物种及其指/趾发育的软骨染色图。
研究团队通过层级聚类分析,将鸟类单个指/趾的转录组与五趾爬行动物(鳄鱼和鳖)的对应指/趾进行比对。结果不仅确认了翼指的1、3、4身份(验证了框架位移),还首次发现了后肢中也存在框架位移:三种鸟类(鸵鸟除外)后肢第二趾均与爬行动物第一趾聚类,提示前肢和后肢中均发生了位置身份与发育身份的错位。
更有意思的是,研究团队提出并验证了一个新假说:鸟类演化出了专性双足行走(obligate bipedalism)之后,前肢从承重的功能性束缚中解放出来,前后肢之间的发育约束随之放松,从而可以各自独立演化出新的功能,就像人类的手和脚一样。数据显示,与四足行走的爬行动物相比,鸟类前肢和后肢之间对应指/趾的差异表达基因数量增加了3到9倍,且这种差异在从肢芽到指/趾发育的过程中持续扩大,在所有鸟类第一指/趾中尤为显著。翅膀和脚趾,就此踏上了各自独立的功能特化之路。
一个源自颌类脊椎动物祖先的古老基因网络
通过定义每根指/趾的“分子指纹基因”(即在特定指/趾中偏好性上调或下调的基因),研究团队将这些基因映射到四足动物系统发育树上,得到了一幅跨越数亿年的演化图谱:
大多数指/趾都具有高度保守的分子身份——这与部分早期研究认为指/趾之间没有固定分子身份的结论相悖。更关键的是,许多维持指/趾前后极性的基因——如Alx4、Pax9、Zic3(前部表达基因,anterior)和Hand2、Hoxd10(后部表达基因,posterior)——在鲨鱼和斑马鱼等鳍芽中就已显示出前后区域的差异表达。这套决定前后轴特化的基因调控网络,可以追溯至有颌类脊椎动物的共同祖先。换言之,数亿年前用于区分鱼鳍前后侧的基因,在今天的四足动物中依然负责区分大拇指和小指。
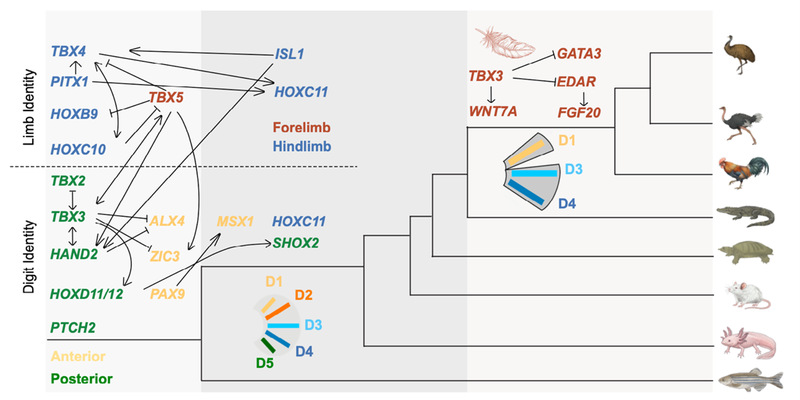
▲ 图二、附肢身份与手指身份的基因网络演化。基于跨物种保守性分析,本研究推断出两套可追溯至有颌类脊椎动物共同祖先的古老基因调控网络: 决定前肢(红色)与后肢(蓝色)的身份以及维持指/趾沿前后轴的分子身份。这些基因差异表达从鱼鳍芽延续至四足动物指/趾阶段。在两套祖先网络的基础上,部分新基因在后续物种分化中逐步加入(四足动物节点如Isl1、Hoxc11;鸟类节点如羽毛发育相关基因),进一步塑造了现生四足动物附肢和指/趾的形态多样性。
从演化蓝图到具体案例:平胸鸟类指/趾的退化和适应
Nature Communications 的论文绘制了四足动物指/趾演化的宏大版图,那么发表在Molecular Biology and Evolution上的研究则像一部深入的特写。研究团队将目光聚焦于平胸鸟类中的两个特殊成员——鸸鹋和鸵鸟,剖析它们翼指退化和脚趾适应性演化的分子细节。
鸸鹋翅膀极度退化,成年个体仅有一根翼指;鸵鸟的后肢也经历了剧烈的重塑:胚胎期有五根脚趾,但在发育过程中第1、2、5趾退化,仅保留第3和第4趾——其中第3趾成为主要的承重趾,高度适应快速长距离奔跑。
通过比较鸸鹋、鸵鸟、鸡以及两种五趾爬行动物(暹罗鳄和中华鳖)的指/趾转录组,研究团队鉴定出与指/趾退化或适应性演化相关的候选基因。分析揭示了一个清晰的规律:鸸鹋翅膀的退化主要涉及肌肉发育通路的基因,而鸵鸟脚趾的退化则主要涉及骨骼发育通路的基因。
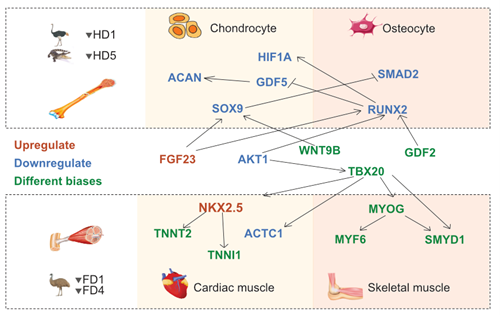
▲ 图三、鸸鹋翼指退化(上图)与鸵鸟/鳄鱼后肢趾退化(下图)由不同的基因调控网络驱动。红色示上调基因,蓝色示下调基因,绿色示在不同手指中调控方向相反的基因。两套网络仅共享极少数基因。
奔跑的利器:鸵鸟承重趾的适应性创新
除了退化,研究团队还捕捉到了一个适应性演化的信号。鸵鸟后肢第3趾作为主要的承重趾,在发育晚期(HH40阶段)表现出独特的分子特征。与骨骼和肌肉发育的基因变化不同,第3趾大量上调了表皮发育相关基因,包括Krt24、Cdkn1a等角质化相关基因以及Pkp1等细胞连接加固基因。这些变化与鸵鸟第3趾已知的更厚表皮和特殊趾垫结构高度一致,为其在高速奔跑中保护软组织、吸收震荡提供了分子层面的解释。
研究团队还追溯到了一个更古老的演化事件。恐龙和现代鸟类的后肢第5趾均退化消失,但鸵鸟和鳄鱼胚胎中仍有残留的软骨结构。通过对两者的比较,研究团队发现约70-77%的退化相关基因在鸵鸟和鳄鱼之间共享,表明第5趾的退化早在主龙类(Archosauria)的共同祖先就已启动,随后在恐龙向鸟类的演化过程中彻底完成。
———————————————————————————————————————
这两项研究一起,讲述了一个关于保守与创新的演化故事:一方面,决定指/趾身份的核心基因网络自颌类脊椎动物祖先以来保持了惊人的深度保守;另一方面,在这套古老网络的约束之上,自然选择可以通过灵活的分子策略——调整不同的发育通路、利用谱系特异的基因表达——来驱动手指的退化、功能重塑和适应性创新。这种底层统一、上层多样的演化逻辑,不仅适用于四足动物的四肢发育,也可能广泛适用于其他同源结构,为我们理解生物表型多样性的起源提供了理论范式。
周琦实验室已毕业博士生康雯为两篇论文的第一作者,周琦教授为两篇论文的通讯作者(其中 Nature Communications与耶鲁大学的 Günter Wagner 教授共同通讯,参与该研究的还有南方医科大学费继锋教授和天津大学的梁聪)。两项研究均受到国家重点研发计划, 国家自然科学基金委,以及浙江大学生命科学研究院的经费支持。
原文链接:https://www.nature.com/articles/s41467-026-73821-7
https://academic.oup.com/mbe/article/43/5/msag101/8654147