线粒体被誉为细胞的“能量工厂”,通过氧化磷酸化为机体生命活动提供绝大部分ATP。然而,线粒体如何根据细胞代谢状态与能量需求,精准调控自身ATP合成能力,其分子机制仍有诸多未解之谜。呼吸链由5个大型复合物协同完成电子传递、质子泵出与ATP合成,其中复合物I包含四十余个蛋白亚基,目前已鉴定出二十多种正向调控其组装的因子,但负向调控机制未见报道【1】。
2026年6月26日,我院佟超实验室在Molecular Cell发表题为 A negative regulator of mitochondrial complex I assembly adapts respiration to cellular energy demand的研究论文,首次报道了定位于线粒体内膜的蛋白TMEM141作为呼吸复合物I组装的负调控因子,能够响应能量需求动态调节氧化磷酸化效率。值得注意的是,虽然TMEM141功能此前未知,但在一个高自杀风险家系中已发现其存在多态性【2】,且在神经发育障碍患儿中也鉴定出该基因突变【3】。本研究不仅填补了呼吸链负调控领域的空白,也为理解TMEM141相关人类疾病提供了新视角。

研究发现,在培养细胞中,当通过改变碳源强制其依赖线粒体氧化磷酸化产能时,线粒体呼吸增强,并伴随TMEM141蛋白水平显著下降。进一步功能实验表明,TMEM141缺失可增强线粒体呼吸并促进复合物I组装,而其过表达则抑制呼吸与复合物I组装,暗示其为复合物I组装的负调控因子。
机制层面,TMEM141定位于线粒体内膜,能够与复合物I组装因子TIMMDC1直接结合,干扰TIMMDC1与亚基NDUFA9的互作,从而削弱TIMMDC1向复合物I组装中间体的掺入,最终抑制成熟复合物I的形成。当细胞能量需求升高、氧化磷酸化水平增强时,线粒体内膜蛋白酶AFG3L2与YME1L1结合TMEM141并介导其降解,从而解除对复合物I组装的抑制,实现呼吸功能的快速上调。
值得注意的是,TMEM141缺失虽能提升能量输出,却同时带来“副作用”:线粒体呼吸过度增强导致活性氧过量产生,进而引发线粒体DNA氧化损伤并释放至细胞质,异常激活cGAS-STING炎症通路。作者通过构建ρ0细胞(缺失线粒体DNA)证实,该炎症通路的激活直接依赖于线粒体DNA释放,提示TMEM141不仅是产能调节器,更是防止线粒体引发免疫过度激活的重要屏障。
在进化保守性方面,果蝇中的TMEM141同源蛋白dTMEM141同样定位于线粒体,其过表达抑制ATP产生并导致线粒体结构异常,缺失则使ATP增加、氧化应激升高,并引起衰老果蝇眼中神经胶质细胞部分丢失,表明该调控机制在进化中高度保守。
综上,该研究首次鉴定TMEM141为线粒体呼吸链复合物I组装的负调控因子,揭示其在基础状态下抑制过度能耗与氧化损伤,在氧化磷酸化需求升高时被降解,从而“解锁”呼吸功能。这一负调控机制为线粒体能量生产提供了关键缓冲,是维持细胞代谢稳态的重要保障。
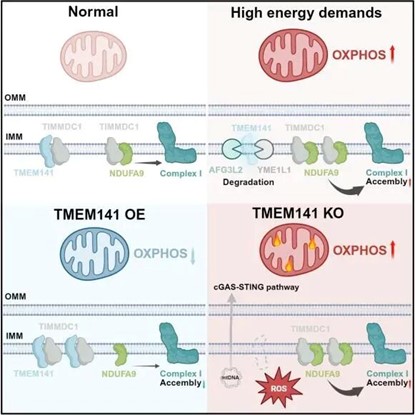
图1 TMEM141调控线粒体氧化磷酸化及细胞稳态
我院佟超教授为论文通讯作者,博士研究生李致融、陈诺为共同第一作者。杭州师范大学衰老研究所刘俊平教授、浙江大学医学院附属第二医院王利权教授对本研究给予大力支持。
参考文献
1. Vinothkumar, K.R., Zhu, J., and Hirst, J. (2014). Architecture of mammalian respiratory complex I. Nature 515, 80-84.
2. M., Callor, W.B., et al. (2013). Genetic risk factors in two Utah pedigrees at high risk for suicide. Transl Psychiatry 3, e325.
3. Sun, L., Yang, X., Khan, A., Yu, X., Zhang, H., Han, S., Habulieti, X., Sun, Y., Wang, R., and Zhang, X. (2024a). Panoramic variation analysis of a family with neurodevelopmental disorders caused by biallelic loss-of-function variants in TMEM141, DDHD2, and LHFPL5. Front Med 18, 81-97.
原文链接:
https://www.sciencedirect.com/science/article/pii/S1097276526003898
实验室主页:
http://lsi.zju.edu.cn/25432/list.htm